Precedence or prescience?
(Much of this was written before the SJI published my article in Sep 2006)
The only honourable justification for claiming precedence of conception (ie, of imagining it before being told of it or reading of it through another author) is that it lends support to the other proposed elements of the hypothesis. Where it was prescient, then this ought to be acknowledged – or it becomes, metaphorically, like Cassandra's tears. The future (direction of understanding) may have been seen but it appears that no one will listen or believe. When something unknown is predicted and found to be supported by the evidence, this is strong support for the hypothesis. I believe that this set of ideas around Morphostasis and the structure that it accords the immune (or morphostatic) system is immensely important. Until the central tenets are understood and accepted by a majority of immunologists, the discipline will continue to fumble around, captive in the vaults of a blind alley. There appears to be continuing, resistance to allowing these ideas to receive an open airing. Many articles are published where citation of the morphostasis articles could be appropriate. It seems unlikely that all authors are completely unaware of the work. Whenever there is discussion of extant theories of what the immune system is about, Cunliffe and Morphostasis are mostly invisible. Perhaps this is because I have done no original research; but that would be a churlish, clubby response. Perhaps it is because people think that I am trying to muscle in on a bandwagon; but that ignores the difficulty I have had getting into print – and when. Perhaps it is because they still see it as far too speculative (and my style to be too naïve) to be "good science"; but, down the "retrospectoscope", it is becoming increasingly clear that – in broad principle – it was prescient. Since very few authors (or journals) will cite the work, it is left to me to emphasise here how it has regularly predicted the way perceptions are about to change. Then, using this historical precedent to extrapolate into the future, much that is still regarded as "sheer speculation" will also probably follow this historical precedent. I believe that many of the ideas are destined to become the respectable "normal science" of the future. Of course, it is important to be cautious and accept nothing without great scepticism (this scepticism is a strength in science). But, it is misguided to believe that mention of it needs to be swept under the carpet to prevent it "corrupting the youth of the realm". Its track record is now far too good. The chance to rapidly apply its manipulative power has been delayed for far too long.
There is clear evidence of tacit and sideways concession that the rough concept has value. Some authors have resurrected earlier and long dormant articles that, in retrospect, made salient (and prescient) points; these seem to establish prior "precedence". However, precedence is irrelevant unless it is paraded to help fan the growing fire of an enlightening perspective that runs counter to extant and accepted perceptions (the latter having a mass action effect). It is the perspective, not any particular author, that should be handed the acclaim; and that demands perceptual readjustment. Personally, I am delighted when I find an old article that was pointing in, what I believe was, the right direction, although it was passed over when published; I have highlighted these when I have found them. I would encourage everyone to look for them and parade them, for they will bring along their own flashes of insight. Suppressing mention of new ideas – because this might be viewed as admitting their possible value is a nonsense and more akin to "religion", in its strict etymological meaning, "religare: to tie up, to tie back".
So what points of "precedence" have I proposed and what do I parade to support them?
- Elective cell suicide is used as a mechanism for defence (this principle can now be easily extrapolated to encompass autophagy, which has more recently emerged as a vital intracellular process).
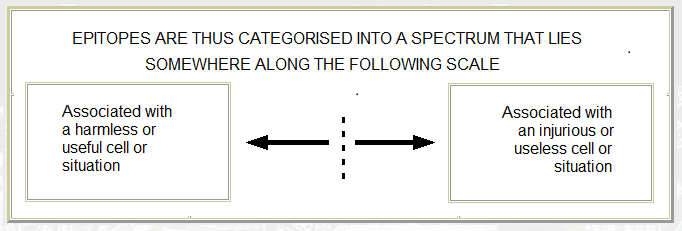
- The primacy and centrality of innate immunity (summarised in Morphostasis and Immunity 1985 – see UNPUBLISHED) and this 1993 diagram
- The adaptive immune system functions in such a way that it acts as a memory for prior innate immune responses (previously encountered pathogenic stimuli) – that is, the system is "inflammocentric".
- The inversion of NK function to Tc function in the vertebrates – linking innate and anamnestic immune systems. There was literature out there to suggest that NK cells attack cells not expressing a self Mhc signature (search Klaus Karre) but I did not discover this until this part of the hypothesis had been established.
- The seamless integration and interdependence of the innate and the anamnestic immune systems (1985 and 1992)
- The conceptual transposition of a lymphocentric to an inflammocentric perspective. This is analogous to the Copernican transposition of the heavenly firmament from being centred about the earth to being centred about the sun.
- The adaptive immune system acts as a memory for inflammation (and thus innate immunity) – it is "inflammocentric". I am not aware of others pressing this perspective. Indeed, there are now a bevvy of articles referring to mechanisms that give innate immunity a memory but they all seem to ignore "the glaringly obvious" point that, in vertebrates, this is the "purpose" of the adaptive system.
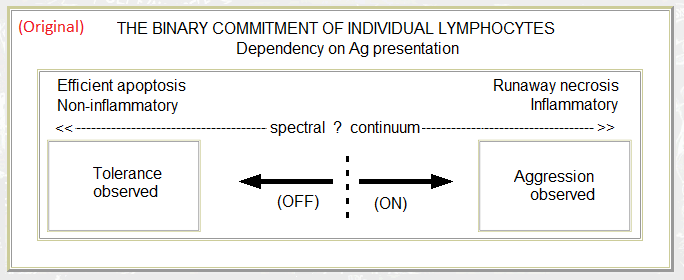
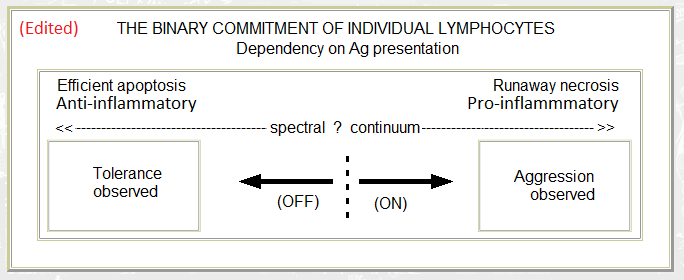
- A "danger" like hypothesis (Morphostasis and Immunity 1985). Personally, I still prefer this version over the "danger" analogy (danger suggesting some pre-existent and anticipable property rather than an after-the-event response to damage). I have swapped the harmless and injurious sides over here so that they now match the second diagram.
- Intracellular surveillance is at the core of the surveillance system (at least as far back as 1994). This is primed ready to encompass "autophagy".
- Plants share the same core defence (morphostatic) mechanism.
- The resurrection of the immune surveillance concept for averting cancer (now creating great excitement). In the late 1980s and early 1990s, immunosurveillance for cancer had been authoritatively condemned to the slag heap by elements of the cognoscenti.
- The polarisation of lymphocyte commitment according to the style of cell death (reproduced from a file I created on 18th July 1993). In retrospect, "Non-inflammatory" and "Inflammatory" may have been better expressed as "Anti-inflammatory" and "Pro-inflammatory". (Various submissions for publication – between July 1993 and the appearance of Polly Matzinger's article in the 1994 Annual Review of Immunology – contained this figure; all were rejected.)
- Gap junctions are important in immune cell interactions.
- Virtually all disease is accompanied by an enhanced auto-rejection and this is roughly synonymous with inflammation (so virtually all disease incurs an inflammatory – phagocyte recruiting – response).
- The realisation that central tolerance is probably the same as peripheral tolerance; it is induced in response to tidy self-cell shutdown in the massive apoptosis that occurs in the thymus. This is looking promising with increasing support for commitment into regulatory T-cells rather than the in utero elimination (death) of all self reactive T-cells.
- And some more – the list is fairly extensive.
- The two "Morphostasis and Immunity" articles (1985 and 1992) are looking increasingly secure. During the years when these ideas remained "obligate heresies" to the reigning cognoscenti, I remember having waves of intense doubt, wondering if they might really present nothing of any value or validity. But, a modern reading returns a different verdict: the (r)ejections – based on the verdict that they were plain wrong and/or unimportant – can now be seen to be unjustified.

The elements that are still "cinderellas" that warrant investigative dissection:
- The involvement of gap junctions. Some of the precise inferences were far too speculative but the liklihood of their actual involvement is high. (Now we need to add tunnelling microtubules into this "recipe".)
- The points raised in "Clinical Morphostasis" – particularly the "two-edged sword" nature of immune activity, where the best response is a Goldilocks "just right" reaction – not too weak and not too strong.
- The points raised in "The neurology of Behçet's syndrome". (These points bring an enhanced perception of the process and emphasise that vasculitis is a manifestation of the intensity of an inflammatory response.)
- The probability that "microbe hunting with the intention of microbe killing" is, at least partly, an illusion and a consequence of the primary function, morphostasis. Morphostasis has evolved from a primitive phagocyte feeding stategy that leads to the management and sequestration of resources that are used for growth and sustenance.
- The adaptive immune system does not "remember" foreign organisms to kill them more quickly on re-encounter. Adaptive immunity does not, by "intention", deploy a "bug" hunting and killing strategy. It does establish a memory of earlier disruptive events (a memory of pathogenic stimuli rather than microbes).
- The shell like system of morphostatic defences. I think that this might prove to be one of the most insightful deductions of them all and, eventually, one of the most useful predictors and explanators. So far, this seems to have been studiously ignored.
- The synthesis outlined in the phlogiston article.
- The obfuscating constriction in the definition of the term pathogen. Definitions range from "any damaging agent" to "a disease causing micro-organism. The second has now become the most frequent interpretation but this is by no justification other than its mass action effect (which is how language drifts). This creates conceptual blinkers that blind us to the "real purpose" of an immune response.
- The possibility/probability that the T-cell repertoire that leaves the thymus consists of (a) committed regulatory T-cells that have been tolerised by apoptotic self-cell death and (b) uncommitted T-cells that can subsequently be channelled into tolerance (the outcome that follows apoptotic death/controlled cell shutdown) or into aggression when the peptide debris is presented during an inflammatory/uncontrolled cell shutdown environment. The old view decided that (virtually) all "self" reactive T-cells must be eliminated (virtually because there was a suspicion of some "leakage").
- And some more. As time goes by, the "retrospectroscope" is lending increasing support to more and more; even where the detailed interpretations have proven wide of the mark, the principles were usually sound.
Recently, it has become abundantly clear that there is a stampede towards a new perspective. "Inflammation, regeneration and tissue homeostasis" have become an in vogue, integrated research arena. The PNAS changed its "Immunology" section to "Immunology and inflammation" on the 15th July 2015. There is no obvious evidence that any of this shift in attention has been influenced by my writing. Indeed, why should it be? If it is largely right (closer to the mirage of "the Truth") then there will, inevitably, prove to be multiple routes to its ultimate realisation, formulation and exposition.
However, I can savour the knowledge of having been one of the first – perhaps even the first – to realise the promise and power that a radical change in perspective could bring. That was, at fleeting instants, orgasmicly mind blowing and sufficient reward in itself.
Finally – even an inexperienced investigator would be able to quickly establish the reality that my articles and the ideas presented in them have been, effectively, "scooped" (unknowingly or knowingly); and scooped following repeated rejections by various journals (including much damning criticism in the process). Simultaneously and out of exasperation, I was sending out a series of letters to key people because I thought that this was too important to sit on alone. There is no justification for parading this other than to stress, "For heaven's sake, someone, take ALL of this seriously." I am sure it has many warts and much amateur naïveté but it also uncovers seams of "conceptual platinum" that are begging to be "mined". I suspect that the refusal to advertise its existence (and the subsequent gagging of my opinion) has slowed progress towards a much deeper understanding and towards an improved technological manipulation of virtually all disease.
I do not consider that it is my place to speculate about the circumstances that may have led to my contribution being eclipsed; for that would just turn into ugly pique. That is the responsibility of society at large; in particular, historians, philosophers and anthropologists who focus on science may, eventually, find this to be "fertile territory" for analysis.
I would have been very interested to attend this workshop (International workshop on the Danger Model). However, the fact that I discovered it only 17 days before it started – did not augur inclusion, respect or welcome. Perhaps that is appropriate.
In the same vein, Immunological Reviews devoted a whole issue in November 2017 (Vol 280) on "Death, Danger and Immunity". I was resigned, on the basis of past experience, to the inevitability that my papers would not feature in either the discussion or the references. My suspicions were justified.
A 2019 book by Walter Gottleib Land made no mention of my work - as far as I can tell (Land W. G. "Damage-Associated Molecular Patterns in Human Diseases" Volume 1: Injury-Induced Innate Immune Responses ![]()
![]() )
)
"You can lead a horse to water but you can't make it drink."
So, the final comments
I suspect that there is a positive reason that this work remains submerged. This reason is almost certainly strategic; the effect leads, without doubt, to the denial of lending me any overt credit for these ideas. I could, in a tortuous way, envisage a benign reason for this but must assume that a malign reason is possible if not probable. Whatever, it has reduced its impact and influence on a large swathe of researchers. Only time will tell if this was a countercurrent in the progress of scientific thinking and manipulation.
All this brings us back to power. I had no power or appropriate influence to get into print right through 1984 to 1994. I still appear to wield no power in bringing substantial attention to my publications. These are out there, visible but largely ignored. Perhaps that is appropriate. But, I suspect it is not. The people that currently assume the mantle of "inventors/creators" of the new perception have the power to bring attention to my writing and its potential precedence. Do they exercise it? And, if not, why not? This question needs addressing by the scientific community; not by me.
I have noticed that, on occasions, commentators have implied that I have built the "morphostasis" concept around Polly Matzinger's "danger" concept. Should you be tempted to accept this, then I suggest that you look through all the historical elements of the origin of the morphostasis theory. These are touched upon on this particular page and throughout this "www.morphostasis.org.uk" website; the rejection letters are particularly relevant. The "morphostasis" perspective was in an advanced state of formulation before Polly's seminal 1994 paper. So, even though my contribution could be categorised as an expansion on the ideas in the "danger theory", it was not built upon them; nor did I need them. Indeed, I believe that the conception should have been formulated as a "damage" (induced) theory of aggressive adaptive immune activation rather than a "danger" (induced) theory of aggressive adaptive immune activation. And that misses out the whole (and dominant) tolerant aspect of adaptive immunity where the volume of tolerant responses far outweighs the aggressive ones. The danger theory appears to reject self-nonself discrimination. I do not. "Healthy-self-cell" vs "other-than-healthy-self-cell" discrimination is, I believe, at the core of the morphostasis concept. This discrimination is made by every cell in the zgygote-derived-colony. However, I categorically reject the concept of self-nonself-epitope discrimination. Instead I promote "damage" (a pathogenic stimulus) as the trigger for pro-inflammatory adaptive immune responses. Tolerance is the outcome of the disposal of a vast deluge of cells that shut themselves down in a controlled and non-inflammatory manner.
So do other proponents of a new view in immunology know of my work? Well, the answer to that is an absolute yes for a number of key authors. Do they attempt to advertise the existence of my contributions? And, if not, why not? There has to be a positive reason, either antagonistic/malevolent or sympathetic/benevolent. I can tortuously imagine a benevolent strategy but, after this length of time, I must assume that a deliberate and polarised avoidance of acknowledging and advertising its existence is more likely to be malevolent.
Finally (finally)
Here are various letters that have passed forward (mostly) and (occasionally) backward ![]() over the years since 1983. I will add more as I find them.
over the years since 1983. I will add more as I find them.
"Osmosis"
PJ, a (then) senior registrar that I worked under, talked a lot about learning "your trade" by (a metaphorical) osmosis. Simply being surrounded by a group of people with ideas and experience in your chosen discipline ensures that a great deal of your eventual knowledge and expertise are absorbed in a passive sort of way. Now, this is a luxury that I would have treasured in immunology but, sadly, being "sent to Coventry" from the early stages has ensured that anything that I learn is done by grinding through journal contents and following search engines. This metaphorical "osmosis" is something that I would love to have experienced.
Is this the epitaph?
It is an interesting analogy that occurs to me now that the USA's GOP has acquitted Donald Trump of guilt to incite insurrection. This highlights how the retention of power, influence and self interest usurp all other considerations - particularly the logical ones. It is quite clear to me now that logical arguments really carried little sway in my attempt to convince immunologists to question their perceptions. When the change comes (it will, inexorably, come now) it will not be because of any criticisms that I have targeted at extant perceptions. So much so that I am now losing interest in broadcasting refinements of my perception of the process. I am now much less inclined to dedicate time and effort on this fruitless exercise. I really don't think that the accusation of "crankery" holds any weight now. Alternative justifications now need to be found to bolster up resistance.
May I suggest "the not new" argument. At the least that might catalyse progress.
I wonder
I have often wondered why my contributions remain so invisible within the field of immunology and its associated literature. I have even wondered whether there could be an industry of deliberate suppression that started long before I was promoted into print. After all, as I pointed out, I was highlighting a "potential seam of conceptual platinum that was begging to be explored and mined". The potential riches (accreditation of precedent, fame, departmental/national renoun and plain simple financial reward) could be staggering. It could be that we are now in a "platimum vs gold rush" to grab credit and acclaim. I am led to wonder whether I have been trampled over in the silent rush (note how Thomas Kuhn described paradigm shifts as "silent" – they only become noticeable as textbooks are rewritten).
Perhaps the historians and philosophers of science will – eventually – be able to get this into perspective.
And, one final churlish thought: did my communications and unsuccessful submissions percolate through – enough – to influence emerging concepts in immunology. The timeline certainly makes this a possiblity. And, would I have kept silent about other "competing" though complementary ideas?